Archives
- 2026-07
- 2026-06
- 2026-05
- 2026-04
- 2026-03
- 2026-02
- 2026-01
- 2025-12
- 2025-11
- 2025-10
- 2025-09
- 2025-03
- 2025-02
- 2025-01
- 2024-12
- 2024-11
- 2024-10
- 2024-09
- 2024-08
- 2024-07
- 2024-06
- 2024-05
- 2024-04
- 2024-03
- 2024-02
- 2024-01
- 2023-12
- 2023-11
- 2023-10
- 2023-09
- 2023-08
- 2023-07
- 2023-06
- 2023-05
- 2023-04
- 2023-03
- 2023-02
- 2023-01
- 2022-12
- 2022-11
- 2022-10
- 2022-09
- 2022-08
- 2022-07
- 2022-06
- 2022-05
- 2022-04
- 2022-03
- 2022-02
- 2022-01
- 2021-12
- 2021-11
- 2021-10
- 2021-09
- 2021-08
- 2021-07
- 2021-06
- 2021-05
- 2021-04
- 2021-03
- 2021-02
- 2021-01
- 2020-12
- 2020-11
- 2020-10
- 2020-09
- 2020-08
- 2020-07
- 2020-06
- 2020-05
- 2020-04
- 2020-03
- 2020-02
- 2020-01
- 2019-12
- 2019-11
- 2019-10
- 2019-09
- 2019-08
- 2019-07
- 2019-06
- 2019-05
- 2019-04
- 2018-11
- 2018-10
- 2018-07
-
ATP binding cassette transporters such as ABCA
2021-06-23
ATP-binding cassette transporters such as ABCA1 and ABCG1 are expressed on cell membranes to transport cholesterol from mst2 to HDL. These transporters help to improve RCT and reduce atherosclerosis. ABCA1 and ABCG1 gene expression are primarily induced by the stimulation of liver X receptor/retino
-
br Experimental procedures br Results
2021-06-23
Experimental procedures Results Fluorescence attributable to GnRH-eGFP neurons was observed through the diagonal band of Broca, to the POA of the hypothalamus in GnRH-eGFP transgenic rats for all groups as described previously (Kato et al., 2003, Tada et al., 2013). Discussion The mEPSCs w
-
br EBV BILF A Virus Encoded TM
2021-06-23
EBV-BILF1—A Virus-Encoded 7TM Receptor with Immune Evasive Functions The EBV-encoded BILF1 receptor (EBV-BILF1) is thought to be implicated in the immune evasion strategy of EBV.56, 61, 62 This orphan 7TM receptor is expressed at significant levels during the early lytic phase of the virus infect
-
br Materials and methods br Results
2021-06-22
Materials and methods Results Discussion Structural analysis of dnmt3 sequences of different ploidy fish cloned in this study showed high similarities to zebrafish sequences, which also contained a unique N-terminal motif and a calponin homology (CH) domain, a conserved proline-tryptophan-t
-
Of note BRQ and LFM are two well
2021-06-22
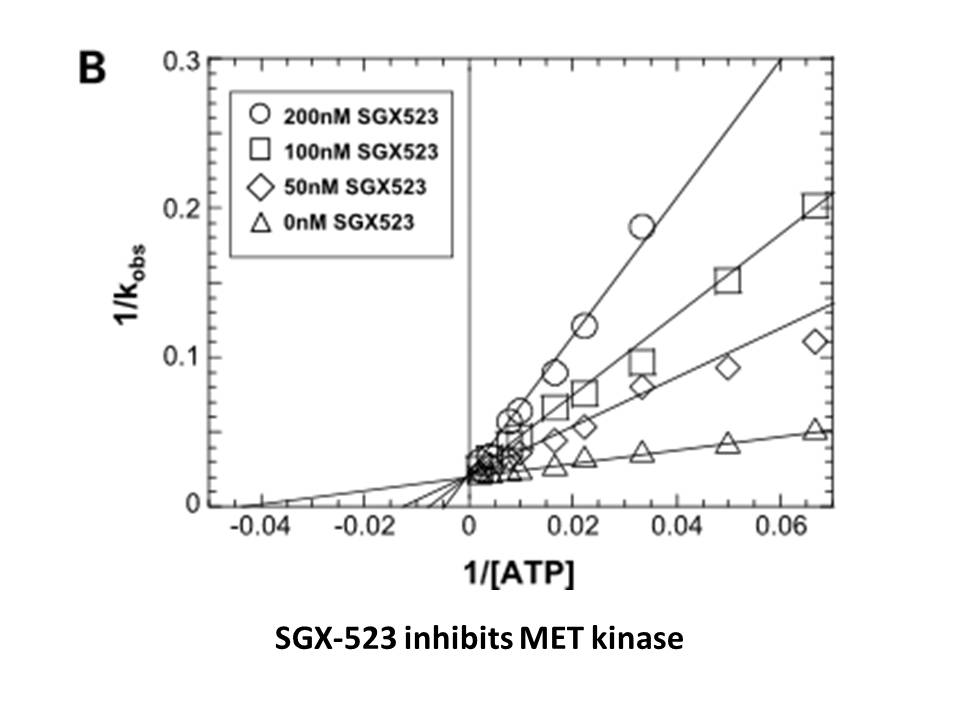
Of note, BRQ and LFM are two well-known immunosuppressive agents (Batt, 1999). As for LFM, it was reported as immunosuppressive therapy for bone marrow (Avery et al., 2004) and renal (Chon and Josephson, 2011) transplantation. Accumulating evidence indicated that organ transplant patients irrelevant
-
br Other Alterations in Triglyceride Metabolism in DGAT Defi
2021-06-22
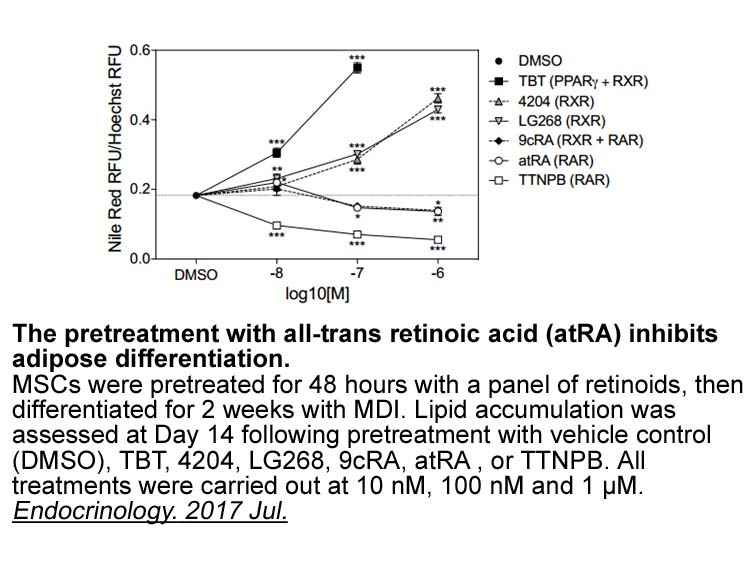
Other Alterations in Triglyceride Metabolism in DGAT-Deficient Mice Consistent with the enzyme's ubiquitous expression pattern, DGAT deficiency also altered triglyceride metabolism in other tissues. For example, postpartum Dgat−/− female mice were unable to nurse their young owing to an absence o
-
br The role of TNFs and AD The TNF superfamily
2021-06-22
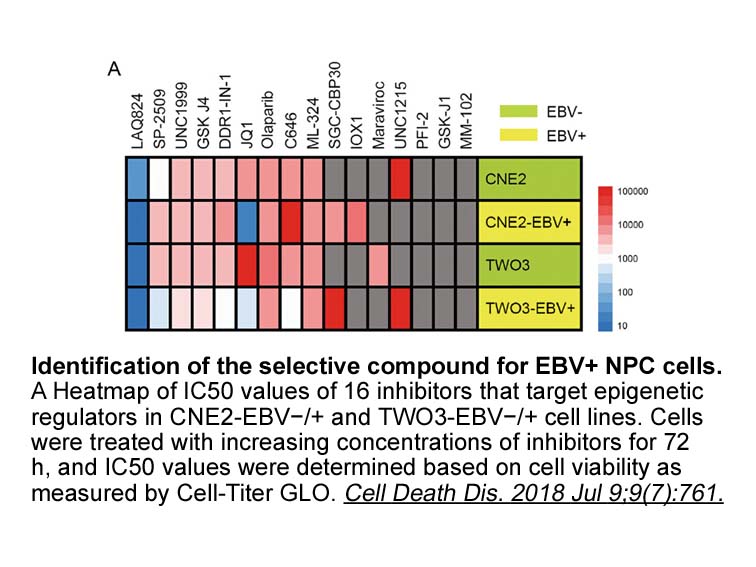
The role of TNFs and AD The TNF superfamily includes 19 members that can bind 29 different receptors [56, 57]; the 19 members are Lta, TNF-α, lymphotoxin-β (LTβ), OX40L, TNF associated activation protein (TRAP, also named CD40L or gp39), factor associated suicide ligand (FasL), CD70, CD153, 4-1BB
-
Furthermore we found that expression of two C
2021-06-22
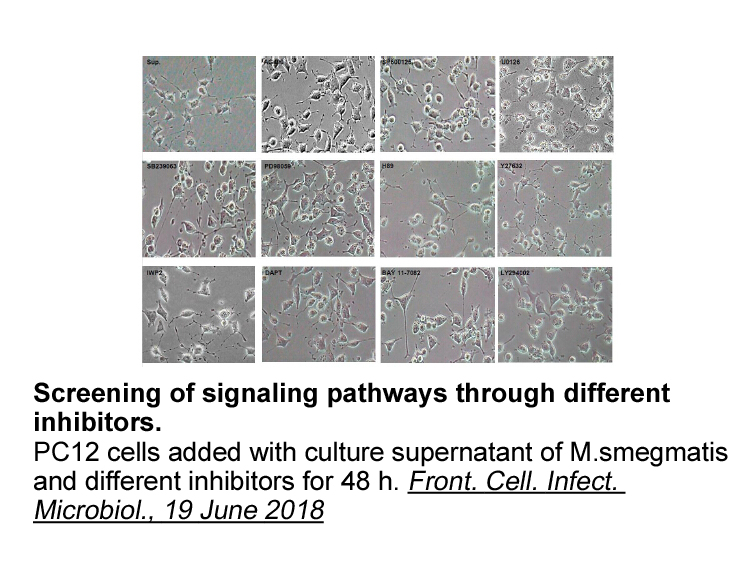
Furthermore we found that expression of two C1 nodule cysteine proteases, Glyma.06G283100 and Glyma.06G174800, changed very little under drought despite the two proteases having similarity to the Arabidopsis senescence-related SAG12 gene with 62% similarity by Glyma.06G283010 and 58% by Glyma.06G174
-
br Conclusion The results of the
2021-06-22
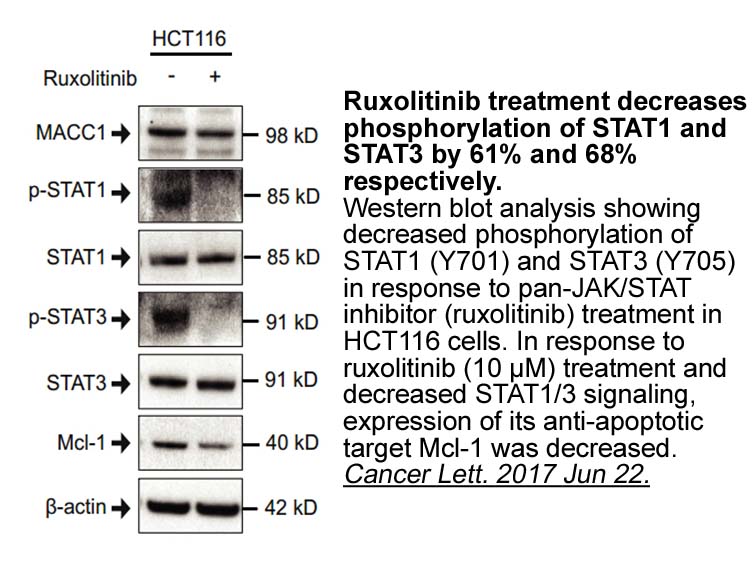
Conclusion The results of the present study demonstrate that cysteinyl leukotrienes LTC4, LTD4 and LTE4 cause moderate to marked whereas the dihydroxy leukotriene LTB4 causes small muscle contraction in the stomach in vitro. The LTD4-induced contraction is mediated by CysLT1 in fundus but by CysL
-
br Conclusions The present study demonstrated that
2021-06-22
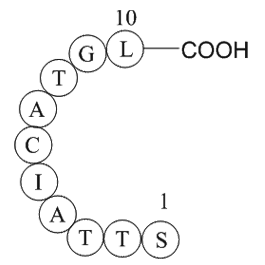
Conclusions The present study demonstrated that CuE possesses strong hepatotoxicity. CuE is not only a mechanism-based inhibitor of human CYP3A4, but also inhibits P-gp activity in vitro. In whole animal studies, CuE induces CYP3A and P-gp after a long-term treatment but inhibits the activities o
-
Increased knowledge about CRC pathogenesis has provided new
2021-06-22
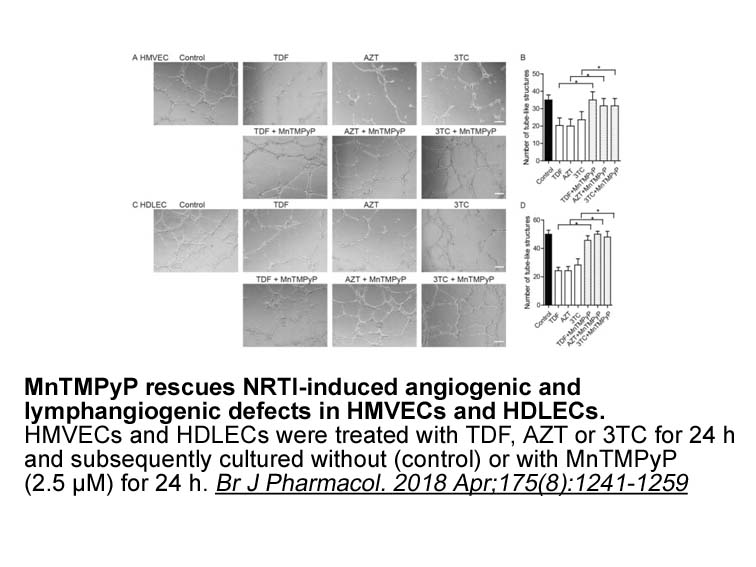
Increased knowledge about CRC pathogenesis has provided new and remarkable treatments beyond the classical options of surgery, radiation and chemotherapy [11]. Despite these advances, chemotherapy remains the most important element of anti-cancer therapy. Chemotherapy, when initiated, is generally e
-
br Introduction Benzanthrone BNZ is a dye intermediate used
2021-06-22
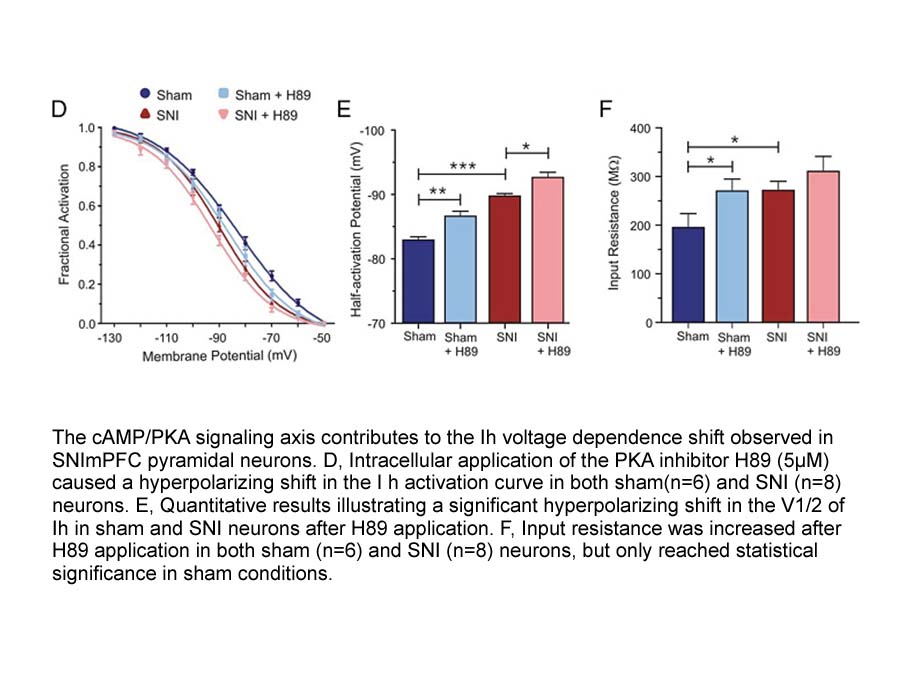
Introduction Benzanthrone (BNZ) is a dye intermediate used in the synthesis of number of polycyclic vat and disperse dyes. It has been detected as an environmental pollutant in urban ambient air particulates, originating from furnace effluents, municipal refuge, wood and coal combustion and autom
-
Today many strategies have been developed to enhance CXCR re
2021-06-22
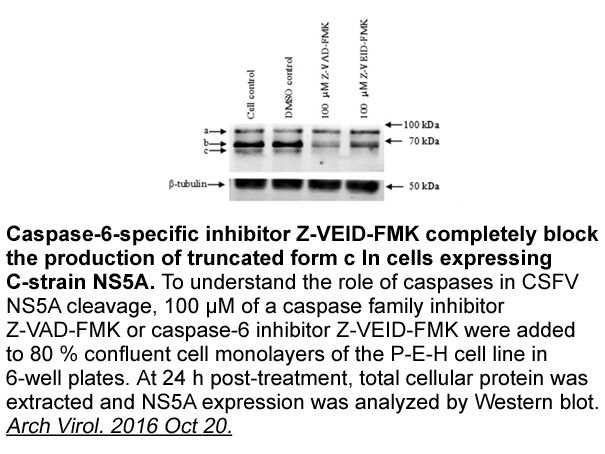
Today, many strategies have been developed to enhance CXCR-4 receptor cell adhesion molecules on the cell surface, either by modifying the cell culture medium or by modifying the CXCR-4 gene in BMSCs to stimulate stem cell recruitment (Jiang et al., 2012, Liu et al., 2014b). For instance, the pre-c
-
The identification of CRF receptor subtypes has led
2021-06-22

The identification of CRF serine protease inhibitors subtypes has led to a number of studies addressing its functional properties. This paper examines the evidence for CRF1 and CRF2 mediation of fear and anxiety. Recent reviews discussing specific CRF receptor functions associated with other condit
-
Fatty acid oxidation plays a crucial role in maintaining bod
2021-06-22
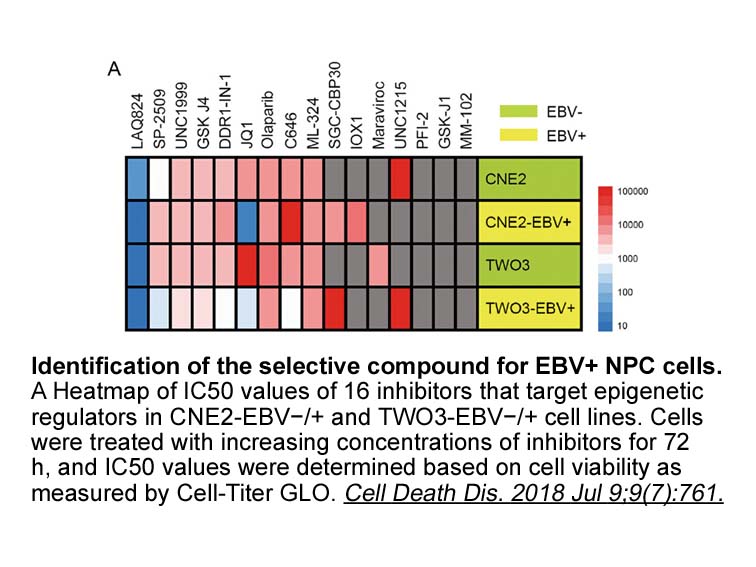
Fatty Biocytin β-oxidation plays a crucial role in maintaining body energy homoeostasis mainly during catabolic states. It mainly occurs in mitochondria and involves a cyclical series of reactions that result in the shortening of fatty acids. Fatty acids comprise an efficient source of energy that r
16581 records 668/1106 page Previous Next First page 上5页 666667668669670 下5页 Last page